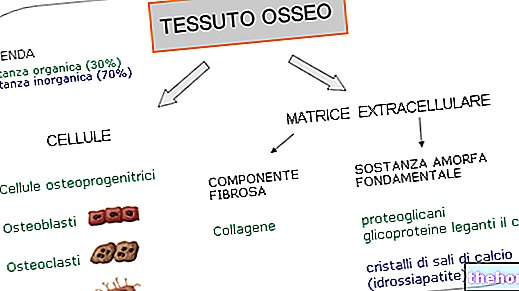
A zsírsavak szintézise az acetil -koenzim A -ból indul, és nagyjából megegyezik a lebomlásuk fordított útjával; a zsírsavak szintézisében a kiindulási acetil -koenzimhez bikarbonát -töredékek sorozata kerül.
A zsírsavak szintézise teljesen citoplazmatikus (azaz az enzimek, amelyek ezt a szintézist katalizálják, megtalálhatók a citoplazmában). A citoplazmában a zsírsavak szintéziséhez használt acetil -koenzim A mitokondriális eredetű: kis része a karnitinon keresztül kerül szállításra, két acil -transzferáz enzim (egy citoplazmatikus és egy mitokondriális) és egy transzlokáz enzim hatására. A mitokondriális eredetű koenzim A -t speciális úton jutják el: a citrát -láz (a név ennek az útnak az első enziméből származik).
A mitokondriumokban jelen lévő acetil -koenzim A piruvát -dehidrogenáz hatását követő glikolízisből származik; Az acetil -koenzim A citrát -szintetáz enzim hatására megy végbe: ez az enzim katalizálja a citrátképződést az acetil -koenzim A és az oxaloacetát reakciójával.Ha a Krebs -ciklus képes kielégíteni az energiaszükségletet, akkor a citrát egy része (a Krebs -ciklushoz felesleges mennyiség) elhagyhatja a mitokondriumokat, és elérheti a citoplazmát, ahol a citrát -láz enzim energiát fordítva visszaváltja acetil -koenzim -A -vé Ily módon lehetséges, hogy az acetil -koenzim A rendelkezésre álljon a citoplazmában, de a képződött oxaloacetátot vissza kell juttatni a mitokondriumokba, hogy újra hozzáférhető legyen a citrát -szintáz enzim számára.
Az oxaloacetát ezután az enzim hatására maláttá alakul malát -dehidrogenáz citoplazmatikus (citoplazmatikus NADH-t használnak): a malát áteresztő metabolit, és újra beléphet a mitokondriumokba, ahol a mitokondriális malát-dehidrogenáz enzim hatására oxaloacetáttá alakul át (NADH-t is kap); a citoplazmatikus páciens alternatívaként alávethető az alma enzimnek, amely dekarboxilezést és dehidrogénezést végez, és piruváttá alakítható. Az alma enzim a NADP + -on működik (hasonló a nikotinamid -adenindinukleotidhoz, de ettől eltérően a két hidroxilcsoport foszforcsoportja a két ribóz egység egyikén), ezért a malátból a piruvátba történő átmenet során NADPH képződik ( amelyet a bioszintézis során használnak) A piruvát ezután belép a mitokondriumokba, ahol a piruvát -karboxiláz hatására oxaloacetáttá vagy a piruvát -dehidrogenáz révén acetil -koenzim A -vé alakul.
Lássunk egy példát: nyolc molekula acetil -koenzim A szükséges a palmitinsav (tizenhat szénatomos lánc) szintéziséhez, de ezek közül csak egyet használnak fel: hét acetil -koenzim -A molekulát az "enzim" acetil -koenzim A karboxiláz (ez az enzim CO2 -molekulát használ, és kofaktorként biotint tartalmaz).
Az acetil -koenzim A karboxiláz enzim szinte inaktív diszpergált formában és aktív aggregált formában (körülbelül húsz egység) létezhet; az átmenet a diszpergált állapotból az aggregátum formába akkor következik be, amikor a citoplazmában "magas citrátkoncentráció van: a citrát" az acetil -koenzim A karboxiláz enzim pozitív modulátora.
Az acetil -koenzim A karboxiláz enzim más pozitív (inzulin) és negatív (glukagon, adrenalin és acil -koenzim A) modulátorokkal is rendelkezik.
Elemezzük a zsírsavak szintézisét az escherichia coli baktériumban, amelyben ez a szintézis hét különböző fehérje hatására megy végbe; az eukarióta sejtekben a zsírsavak szintézisének mechanizmusa hasonló a baktériumokéhoz, de az eukariótákban a szintézisért felelős hét enzim két multienzim A és B komplexbe van csoportosítva.
A baktériumokban hét különböző gén kódolja:
- ACP (acil hordozó fehérje);
- ACP-acetil-transzacetiláz;
- ACP. Malonil -transzetiláz;
- β-keto-acil-ACP szintáz (kondenzáló enzim);
- β-keto-acil-ACP reduktáz;
- D-p-hidroxi-acil-dehidratáz;
- enoil-ACP szerkesztve.
Az eukariótákban két gén kódolja:
A alegység
AKCS;
Kondenzáló enzim
β-keto-acil-ACP reduktáz.
B. alegység
ACP-acetil-transzacetiláz;
ACP-malonil-transzetiláz;
D-p-hidroxi-acil-dehidratáz;
enoil-ACP szerkesztve.
Az Escherichia coli hét fehérje úgy van elrendezve, hogy van egy központi (az AKCS), és a másik hat körülötte.
Két szulfhidrilcsoport vesz részt enzimatikus hatásában: az egyik egy ciszteinhez, a másik egy foszfopantetin hosszú karjához tartozik; Az ACP kötődik a szubsztrátumhoz, amely a foszfopantetin karon keresztül érintkezésbe kerül a többi enzimmel, amelyek így képesek elvégezni enzimatikus hatásukat.
Az acetil-koenzim A (ACP-acetil-transziláz segítségével) kötődik az ACP-enzimhez (pontosabban a cisztein-kénhez, amely a ciszteil-származékot képezi), és felszabadul a koenzim; az ACP-malonil-transziláz beavatkozik, ami katalizálja a malonil foszfopantethein -en (ebben a folyamatban az eredetileg malonilhoz kötődő A -koenzim is felszabadul).
A következő lépés a β-keto-acil-ACP szintáz, amely egy kondenzáló enzim: lehetővé teszi a két csontváz közötti fúziót; A malonil könnyen dekarboxilálható, és az acetil-származék cisztein karbonilcsoportja képződik: a cisztein felszabadul, és β-keto (acetil-acetil) -foszfopantetin-származék képződik.
Ezt követően a β-keto-acil-ACP reduktáz beavatkozik, amely a karbonilt tovább redukálja az ACP-enzimhez (a NADPH hidroxidot képez, amely NADP +-ra redukálódik).
Most a 3-hidroxi-acil-ACP dehidrátáz hat (dehidráció következik be), ami telítetlen rendszer (alkén) kialakulásához vezet.
A következő folyamat az enoil-ACP-reduktázt tartalmazza (hidrogénezést hajt végre: az alkán képződik, és a NADPH NADP +-ra redukálódik).
Az utolsó fázis magában foglalja az első ciklusból kapott aciltermék átalakítását olyan vegyületté, amely képes egy második ciklus elindítására: a transaciláz enzim átviszi az acilt a ciszteinre, szabadon hagyva a pantetin helyét, amely most hajlandó lesz megkötni egy másik ciklust malonyl.
A β-oxidáció során egy FAD molekulát használnak fel a telítetlen α-β metabolit transz-enoil-koenzim A dehidrogénezéssel történő előállítására; a zsírsavak szintézisében ehelyett egy NADPH molekulát használnak az ellenkező reakció kiváltására.
Általában tizenhat szénatomos zsírsavakat szintetizálnak, de tizennyolc, húsz vagy huszonkét szénatomot tartalmazó zsírsavak is előállíthatók; a zsírsavakat ezután észterezik, és aktivált glicerinnel (azaz glicerin-3-foszfáttal) triglicerideket képeznek. Ez utóbbi dihidroxi -aceton -foszfátból nyerhető az enzim hatására glicerin -foszfát -dehidrogenáz vagy glicerinből az enzimen keresztül glicerin kináz.
A szintetizált zsírsavakat a zsírszövetbe kell küldeni; a véráramban trigliceridek formájában, vagy részben önmagukban transzporterfehérje, azaz albumin segítségével szállítják.

.jpg)
.jpg)