Az élőlények ciklikus folytonossága a szaporodás jelenségeiben találja meg az összefüggéseket az egymást követő generációk között.
A reprodukció az evolúciós skála különböző szintjein, a növény- és állatvilág különböző következményein, a különböző élő fajokon történik, olyan mechanizmusok sokféleségével, amelyek önmagukban egy egész értekezést indokolnak.
A szaporodás jelenségeinek első osztályozásakor meg kell különböztetni az egysejtűeket a többsejtű organizmusoktól, mivel csak az előbbiben esik egybe a sejtosztódás a szaporodással.
A többsejtűekben a szaporodás lehet agamikus vagy szexuális (vagy gamikus).
Az agamikus szaporodás viszonylag ritkán a mitózis mechanizmusán alapul, így a fajok változékonyságát inkább a mutációk ismételt előfordulására bízzák.
Különféle mechanizmusok is léteznek, például strobilizáció, sporuláció stb., Míg a növényekben a mezőgazdaságban jól ismert regenerációs formákat találunk (dugványok, rétegezés stb.).
A magasabb formákban a legelterjedtebb szaporodási mechanizmus azonban a szexuális, amely megfelel a meiózis megjelenésének, az ivarsejtek képződésének és a zigóta fúziójának (megtermékenyítés).
A primitív fajoknál az ivarsejtek morfológiailag nem differenciálódnak: ebben az esetben izogametiáról beszélünk. Mindazonáltal mindig két ivarsejt-sorozat létezik, amelyeket a (+) és (-) szimbólumokkal azonosítanak, és a megtermékenyítés csak az "ellentétes előjelű ivarsejtek közötti találkozásból következhet be: ezért biológiai különbség van, még nem fejeződött ki morfológiailag.
Az evolúciós skála előrehaladtával megjelenik a morfológiai és funkcionális differenciálódás, amelyben a női ivarsejtek egy típusa általában bőséges tartalékanyaggal rendelkezik (a deutoplazma vagy borjú, amely biztosítja az embrió fejlődését mindaddig, amíg metabolikusan független nem lesz), és egyfajta hím ivarsejtek, amelyek mozgékonysággal rendelkeznek, hogy elérjék a nőstényeket.
A meíózis és a zigóta között sorozatos haploid sejtgenerációk múlhatnak el, akárcsak a zigóta és a meiózis között diploid sejtgenerációk sora, a generációs váltakozással kifejezve a generációs ciklusok különböző fajtái.
A diplonte fajra (diploid szervezetre) a gametikus meiózis jellemző: a meiozis közvetlenül termeli az ivarsejteket, amelyek az egyesítéssel azonnal helyreállítják a diploid állapotot. Ez a metazoanok elterjedt esete, beleértve az embert is.
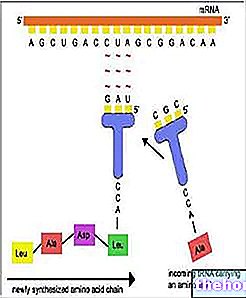
Gametogenezis
Figyelembe véve a gametikus meiózissal történő reprodukciót, ahogy az emberben előfordul, próbáljuk tisztázni, hogy a meiózis hogyan illeszkedik a gametogenezishez (ivarsejtek képződése).
Az embrionális fejlődésben, a hím és nőstény gametogenezisben (spermatogenezis és oogenezis) korai differenciálódás figyelhető meg a testet alkotó (szomatikus vonal) és az ivarsejtek termelésére szánt sejtek (csíravonal) között. A csíravonal kezdeti sejtjeit protogonoknak nevezzük. A nemi nemek férfi vagy női értelemben történő differenciálódásával a csírasejtek differenciálódnak spermatogóniára és ovogóniára.
A spermatogenezist tekintve azt látjuk, hogy a spermatogóniában számos sejtgeneráció van, amely egész életen át folytatódik. Az így folyamatosan termelt spermatogónia csak egy része különbözik a normál mitotikus ciklustól, és a meiotikus ciklus kezdődik helyette.
A csírasejtet, amelyben a meiózis megkezdődik (reduplikáció, majd első osztódás), elsőrendű spermatocitának nevezzük; osztódása két másodrendű spermatocitát eredményez, amelyek a második osztódással összesen négy spermatidát eredményeznek.
A kromoszóma -készlet redukcióját az elsőrendű spermatociták 4n -ből (reduplikáció után négy kromatilátus van minden homológpárra) a második rendű spermatociták 2n -jére és a spermatidák n -jére oszthatjuk, amint azt már tanulmányoztuk meiosis, amelyet így következtetünk. A spermatidák tehát már haploidok, de még nem érett ivarsejtek. A haploid típusú sejtek szerkezetéből a funkcionális érés (az úgynevezett spermiohistogenesis) a spermatidákat spermiumokká, azaz érett hím ivarsejtekké alakítja át.
A női gametogenezisben (vagy oogenezisben) számos különbség van. Először is, az előkészítendő ivarsejtek száma sokkal kisebb. Becslések szerint körülbelül 5x105 ovogon készül az emberi faj nőstényének ivarmirigyében; közülük csak mintegy 400 érdeklődik a tüszőérés és az azt követő dejunktúra iránt, egy olyan ciklusban, amely általában csak havi egy tüszőt érint, körülbelül 35 éves termékeny időszakban.
A két nemben előállított ivarsejtek eltérő száma megfelel a már említett funkció- és viselkedésbeli különbségeknek: a spermiumok kicsik, mobilok és sokfélék a tojás keresésének szükségessége és a találás alacsony valószínűsége miatt; nagy, inert és kevés, az embrió tartalékanyagának garantálása és a belső megtermékenyítés által biztosított védelem szempontjából (természetesen, különösen a külső megtermékenyítésnél a petesejteknek is többnek kell lenniük).
Az ivarsejtek tartalékanyaggal való ellátásának szükségessége megfelel az "oogenezisben" a meiózis leállításának egy szakaszában, amelynek során a kromoszómák részben despiralizálódnak. Ezután megfigyeljük az úgynevezett "toll-kromoszómákat", amelyekben egy sor Az extroverziók azonosítja azokat a tulajdonságokat, amelyekben a deutoplazma szintéziséért felelős gének lecsökkennek.
A nőstény ivarsejtekhez szükséges kisebb szám megfelel annak a ténynek is, hogy a meiózis által termelt négy haploid sejt közül csak az egyik kapja meg a teljes tartalékanyagot, és lesz ivarsejt, míg a másik három (a polociták vagy poláris testek), amelyek csak kromoszóma -anyag, nem hozhat létre zigótákat és embriókat, és visszaszorulnak.
Trágyázás
A megtermékenyítés, vagyis a hím és nőstény ivarsejt találkozása nagyon különböző módon hajtható végre.Az állatvilágban megfigyelhető az átmenet a külső megtermékenyítésről (az ivarsejtek bármilyen környezeti kockázatnak kitett és ezért szükségszerűen nagyon sok a két nemben) a belső megtermékenyítésre, amelyhez a szülői gondoskodás tovább kapcsolódik az emlősök anya és magzat közötti metabolikus kapcsolatához .
A megtermékenyítésnek, miután megtörtént az ellenkező nemű ivarsejtek közötti találkozás, két feltételt kell garantálnia: a sajátosságot és az egyediséget. Vagyis biztosítani kell, hogy a spermiumok azonos fajtájúak legyenek, mint a tojás, és hogy az első belépés után más ne lépjen be.
A specificitást az akroszóma és a petesejt felszínének biokémiai jellemzői biztosítják. Valójában a "termékenyítők" és az "antifertilizinek" közötti reakciókról beszélnek, amelyek specifikussága összehasonlítható az enzimek és a szubsztrát találkozásával.
A megtermékenyítés egyediségét a petesejt felszíni szerkezetének módosítása („kortikális reakció”) garantálja, amely közvetlenül az első specifikus trágyáz / antifertilisin reakció pillanatában kezdődik; e reakció után a petesejt membránja megváltozik, így az azt elérő más spermiumok már nem képesek elindítani a specifikus megtermékenyítési reakciót.
A megtermékenyítést követően a spermiumok farka kívül marad a petesejten, míg a kromoszóma -anyag behatol rajta. Ez, az úgynevezett "hím pronucleus", csatlakozik a petesejt "női pronucleusához", így képezve a zigóta diploid magját.