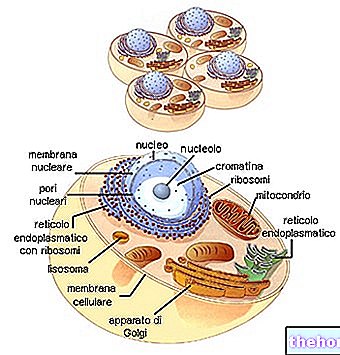
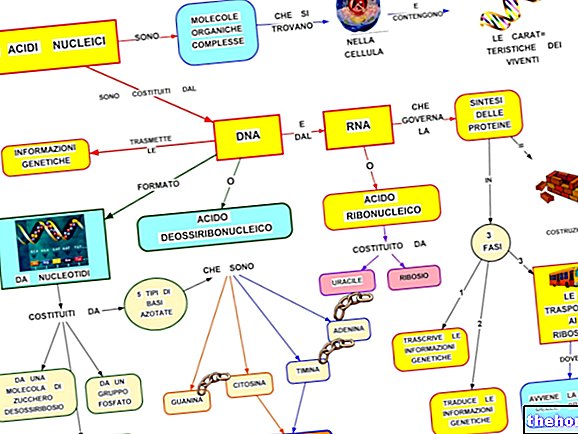
Ahhoz, hogy megfeleljen a polinukleotid és a polipeptid információi között, létezik egy kód: a genetikai kód.
A genetikai kód általános jellemzői a következők:
A genetikai kód hármasokból áll, és nem tartalmaz belső írásjeleket (Crick & Brenner,).
Ezt "nyílt sejtfordító rendszerek" segítségével sikerült megfejteni (Nirenberg & Matthaei, 1961; Nirenberg & Leder, 1964; Korana, 1964).
Nagyon elfajult (szinonimák).
A kódtábla megszervezése nem véletlen.
Hármasok "hülyeség".
A genetikai kód "standard", de nem "univerzális".
A genetikai kód táblázatát tekintve emlékeznünk kell arra, hogy az "RNSm polipeptidre történő lefordítására vonatkozik, amelyben az érintett nukleotidbázisok A, U, G, C. A polipeptidlánc bioszintézise a a nukleotidszekvencia aminosav szekvenciában.

Az RNAm mindegyik bázis triplettjének, amelyet kodonnak neveznek, az első bázis a bal oszlopban van, a második a felső sorban, a harmadik a jobb oszlopban. Vegyük például a triptofánt (azaz Try), és látjuk, hogy a megfelelő kodon legyen rendben, UGG. Valójában az első bázis, U, magában foglalja a tetején található dobozok teljes sorát; ebben a G azonosítja a jobb szélső dobozt és magának a doboznak a negyedik sorát, ahol az írott Try -t találjuk. Hasonlóképpen, a Leucine-Alanine-Arginine-Serína (Leu-Ala-Arg-Ser szimbólumok) tetrapeptid szintetizálásához megtaláljuk az UUA-AUC-AGA-UCA kodonokat.

Ezen a ponton azonban meg kell jegyezni, hogy tetrapeptidünk összes aminosavát (a triptofánnal ellentétben) több kodon kódolja. Nem véletlen, hogy a most közölt példában a jelzett kodonokat választottuk, és ugyanazt a tripeptidet más RNSm-szekvenciával, például CUC-GCC-CGG-UCC kódolhattuk volna.
Kezdetben az a tény, hogy egyetlen aminosav több mint egy hármasnak felelt meg, a véletlenszerűség jelentését kapta, kifejezve a szinonimia jelenségének meghatározására használt kód degenerációjának kifejezésében is. Másrészről egyes adatok arra utalnak, hogy a genetikai információk eltérő stabilitására utaló szinonimák elérhetősége egyáltalán nem véletlen. Ezt látszik megerősíteni az A + T / G + C arány eltérő értékének megállapítása is az evolúció különböző szakaszaiban. Például a prokariótákban, ahol a változékonyság igényét nem elégítik ki a mendelizmus és a neo-mendelizmus szabályai, az A + T / G + C arány hajlamos a növekedésre. Az ebből adódó alacsonyabb stabilitás a mutációkkal szemben nagyobb lehetőségek a génmutációból véletlenszerű változékonyságra.
Az eukariótákban, különösen a többsejtű sejtekben, amelyekben szükség van arra, hogy az egyetlen szervezet sejtjei ugyanazt az örökletes örökséget megőrizzék, a DNS A + T / G + C aránya csökkenni fog, csökkentve a szomatikus génmutációk lehetőségét .
A szinonim kodonok megléte a genetikai kódban felveti a már említett problémát az antikodonok sokaságával vagy sem az RNSt -ben.
Bizonyos, hogy minden aminosavhoz legalább egy RNSt tartozik, de nem egyformán biztos, hogy egyetlen RNSt képes -e kötődni egyetlen kodonhoz, vagy közömbösen felismeri a szinonimákat (különösen akkor, ha ezek csak a harmadik bázis esetében különböznek egymástól).
Azt a következtetést vonhatjuk le, hogy minden aminosavhoz átlagosan három szinonim kodon tartozik, míg az antikodonok legalább egy, de legfeljebb három.
Emlékeztetve arra, hogy a géneket a DNS nagyon hosszú polinukleotidszekvenciáinak egyetlen szakaszának szánják, egyértelmű, hogy az egyetlen gén elejét és végét szükségszerűen a memóriában kell tartani.
A fehérjék bioszintézise
A DNS különböző részeiben ott van a kettős lánc megnyitása és a különböző típusú RNS szintézise.
A betöltési lépés során az RNSt az aminosavakhoz kötődik (amelyeket korábban az ATP és a specifikus enzim aktivált). A bioszintetikus "gépezet" nem képes "kijavítani" a helytelenül betöltött tRNS -eket.
Az RNSr ezután a két alegységre osztódik, és a riboszómális fehérjékhez kötődve a riboszómák összeállását eredményezi.
A citoplazmán áthaladó RNSm a riboszómákhoz kötődve létrehozza a poliszómát.Minden egyes, a hírvivőn áramló riboszóma fokozatosan befogadja a relatív kodonokkal komplementer RNSt -t, felveszi az aminosavakat és megköti őket a polipeptidlánchoz.
A viszonylag stabil RNSt újra belép a keringésbe. A riboszómákat is újra felhasználják, felszabadítva a már összeállított polipeptidet.
A hírvivő, kevésbé stabil, mivel minden egykatatén, (a ribonukleáz által) hasad az alkotó ribonukleotidokká.
A ciklus így folytatódik, és egymás után szintetizálja a polipeptideket a transzkripció által szállított hírvivő RNS -eken.