Ha egy sejt anaerob körülmények között dolgozik, akkor energiát termel a glükóz laktáttá alakításával, és a Cori -cikluson keresztül elhasználja azt; ha rendelkezésre áll oxigén (ezért nyugalmi körülmények között), a glükóz több mint 90% -a aerob módon kerül felhasználásra. csak a maradék 10%, anaerob módon. Ha több ATP -re van szükség, mint amennyit az aerob útvonal képes biztosítani (például amikor az izmok stressz alatt vannak), akkor a kiegészítő ellátást az anaerob anyagcsere biztosítja (oxigénhiányos állapotokban vagyunk: légszomj, fáradtság) légzésben stb.): fel kell gyorsítani ezt az anyagcserét azáltal, hogy a laktátot (amelyet glikolízis útján nyernek) glükózzá alakítanak glükoneogenezis útján.
Aerob anyagcsere alakul ki a mitokondriumokban.
Az első enzim, amely az aerob anyagcserében előfordult, az piruvát -dehidrogenáz; Pontosabb, ha azt mondjuk, hogy a piruvát-dehidrogenáz enzimkomplex, nem pedig enzim, mivel 48-60 fehérjeegység aggregátuma, és három katalitikus hely működik egymás után.
A piruvát -dehidrogenáz katalizálja a következő reakciót (redox):
Piruvát + NAD + + CoA-SH → Acetil-CoA + NADH + H + + CO2
A CoA-SH koenzim A: a pantoténsav származéka; az acetil-koenzim A tioészter. Ez egy redox folyamat, mert a piruvát első szénje a harmadik oxidációs szintről a negyedik oxidációs számra (oxidálódott), a második piruvát szén pedig a második számú oxidációról a harmadik oxidációs számra (ez oxidálódott). Ezután a piruvát oxidálódik (összesen két elektronot veszít), és a NAD redukálódik.
Mint említettük, a piruvát -dehidrogenáznak háromféle enzimatikus aktivitása van, mindegyiket saját katalitikus kofaktoruk támogatja:
- tiamin -pirofoszfát (a B1 -vitamin származéka); deprotonált formában aktív: karbanion képződik.
- lipoamid (a liponsav származéka); nagyon reakcióképes diszulfid hidat tartalmaz.
- flavin -adenin -dinukleotid (a B2 -vitamin származéka); redox tulajdonságokkal rendelkező nukleotid: redoxközpontja a flavinból áll.
Az eukarióta sejtekben az aerob anyagcsere a sejt speciális organelláiban történik, amelyek a mitokondriumok; a baktériumokban a glükóz és más fajok metabolizmusa a sejtben történik, de nincsenek speciális organellák.
Amikor a piruvát belép a mitokondriumba, "piruvát -karboxiláz hatásának van kitéve, ha glükoneogenezisre van szükség (a kiindulási anyag rekonstruálásához), vagy ha piruvát -dehidrogenáznak van kitéve, ha energiatermelésre van szükség: "Az aerob anyagcserével keletkező acetil -koenzim A stimulálja a piruvát -karboxiláz hatását, ezért elősegíti a glukoneogenezist és csökkenti a piruvát -dehidrogenáz hatását.
Nézzük most, hogyan működik a piruvát -dehidrogenáz; mindenekelőtt a piruvát tiamin -pirofoszfát hatására dekarboxileződik.
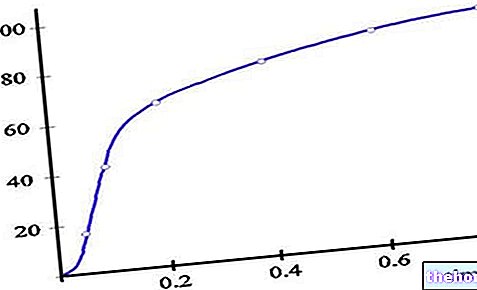
A savas környezet gátolhatja az aerob anyagcserét, mivel a tiamin -pirofoszfát anionos formája aktív, amely savas pH -n protonálódna, és dekarboxilezés nem történne meg.
A dekarboxilezés nehéz reakció, mivel a szén-szén kötést meg kell szakítani; ebben az esetben a reakciót termodinamikailag előnyös az a tény, hogy a reakcióközbenső termék (hidroxietil-tiamin-pirofoszfát) rezonanciát ad (a molekula p-elektronjai delokalizálódnak): a hidroxi-etil-tiamin-pirofoszfát három lehetséges formában (rezonancia) létezik, és ez eléggé stabillá teszi. Továbbá, a hidroxi-etil-tiamin-pirofoszfát anionos formában elég hosszú ideig fennmarad ahhoz, hogy kölcsönhatásba léphessen a lipoamid diszulfidhídjával (a piruvát-dehidrogenáz második katalitikus kofaktorával); a diszulfid-híd egy oszcilláló kar (a hosszú rugalmas lánc vége), és az enzimkomplex egyik katalitikus helyéről a másikra mozoghat.
Ezután a lipoamid a diszulfidhídon keresztül megköti a hidroxi-etil-tiamin-pirofoszfátot: acetil-lipoamidot kapunk. Ez az első fázis a transzcetilezési reakcióban, amelyet a piruvát-dehidrogenáz komplex első enzime katalizál; ebben a fázisban megszakadt a kötés a hidroxilcsoport és a tiamin -pirofoszfát, amelyek eredeti formájukba tértek vissza: redox reakció zajlott le, amelyben a diszulfid -híd oxidálószerként (a két kénatom redukálva) hatott a hidroxilcsoport felé, amelyet acetilré oxidált.
E fázis után a lipoamid oszcilláló karja elmozdul és közeledik a piruvát -dehidrogenáz második enziméhez, amely az acetilcsoport hordozásával végzi el az igazi transzacetiláz -aktivitást: a második enzim által katalizált transzacetilezési reakció második fázisa következik be; Ily módon acetil -koenzim A -t kaptunk. Most helyre kell állítani a redukált formában lévő lipoamidot: a piruvát -dehidrogenáz harmadik enzime lép közbe, amely redoxálja a lipoamidot, és átviszi elektródáit a FAD -ra, amely FADH2 -re redukálódik. A FAD / FADH2 redoxpárként funkcionálhat két különálló monoelektronikai szakaszban vagy egyetlen bielektronikus szakaszban.
A FADH2 azonnal átadja elektronjait a NAD + -nak, és megkapja a FAD -t és a NADH + H + -t.
A leírtak szerint előállított acetil -koenzim A a Krebs -ciklus (vagy a trikarbonsav -ciklus) kiindulási terméke.