Mendel, Gregor - cseh természettudós (Heinzendorf, Szilézia, 1822 -Brno, Morvaország, 1884). Ágoston -rendi testvérré lépve belépett a brnói kolostorba 1843 -ban; Ezt követően a bécsi egyetemen végezte tudományos tanulmányait, 1854 -től Brnóban fizikát és természettudományokat tanított. 1857 és 1868 között hosszú gyakorlati kísérleteknek szentelte magát a borsó hibridizációjával kapcsolatban a kolostorkertben. Az eredmények gondos és türelmes megfigyelése után világos és matematikai pontossággal megfogalmazta a fontos törvényeket, amelyek Mendel törvényei néven szerepelnek. Ezek a törvények ugyanúgy érvényesek a növényvilágra, mint az állatokra, és ezek a biológiai tudományok új ága, a genetika kiindulópontját jelentették. Kilenc évig, száz és száz mesterséges beporzás eredményeit elemezve, mintegy 12 000 növényt művelve és vizsgálva, Mendel türelmesen rögzítette minden észrevételét, amelyek eredményeit 1865 -ben a Brnoi Természettudományi Társaság rövid memoárjában ismertették. a kiadványt nem értékelték minden jelentőségében, és nem keltette fel az "érdeklődést, amit megérdemelt. A tudósok több mint harminc éve" figyelmen kívül hagyták, a törvényeket 1900 -ban egyidejűleg és függetlenül fedezte fel három botanikus: H. de Vries Hollandiában , C. Currens Németországban, E. von Tschermak Ausztriában; de időközben a biológia tanulmányozása nagy előrelépést tett, az idők megváltoztak, és a felfedezés azonnal nagy hatást gyakorolt.
Az első törvényt, vagy az erőfölény törvényét helyesebben a hibridek egységességének törvényének is nevezik. Mendel két borsónövényt vett (amelyeket ő őseinek nevezett), mindkettő tiszta fajta, az egyik sárga, a másik zöld, és az egyik pollenjét felhasználta a másik megtermékenyítésére. Ebből a keresztből származik a hibrid növények borsójának első generációja, amely már nem tiszta nemesítésű; minden növény sárga magvú borsót termelt, egyik sem mutatta a zöld magvú jelleget. Más szóval a sárga betűtípus uralta a zöldet; vagyis a sárga volt domináns, zöld, maszkos, recesszív. Van egy különleges eset is, amikor hiányos dominancia van, és az első generáció köztes jelleget mutat az apai és az anyai között; de még ebben az esetben is egyenlőek lesznek a hibridek egymással. Mendel ragyogó és ötletes magyarázatot adott a jelenségekre; feltételezte, hogy az ivarsejtekkel együtt a karakterek fejlődéséért felelős tényezőket is továbbítják; úgy gondolta, hogy minden szervezetben vagy egy adott karakterben két tényező szabályozza, az egyiket az anya, a másikat az apa közvetíti, és hogy ez a két tényező azonos a fajtatiszta egyedeknél, más a hibrideknél, és hogy végül csak egy tényező szerepel ivarsejtekben. Mendel az antagonista karakterek két tényezőjét az ábécé betűivel jelölte, nagybetűvel az uralkodó, kisbetűvel a recesszívvel; és mivel minden szülőnek van néhány tényezője, például AA -val jelezte a borsót, amely a domináns sárga karaktert hordozza, a hibrid, amely a zöld recesszív karaktert hordozza.
Itt rámutathatunk arra, hogy az egyed megjelenése alapján nem mindig lehet tudni, hogy tiszta fajtához tartozik -e, vagy hibrid, ehelyett meg kell vizsgálni a viselkedését a keresztezésben és a keresztezésben. Valójában a fajtatiszta sárgaborsó és a hibrid borsó látszólag azonos; azonban ismert, hogy genetikai összetételük eltérő, az egyik AA, a másik Aa. Míg a tiszta fajtájú sárgaborsót (AA) keresztezi, mindig és csak sárga magvú borsója lesz, a sárga vagy félig sárga, de hibrid borsó (Aa) keresztezésével pedig zöld magvakkal rendelkező növények is megjelennek utódaikban . A sárgaborsó Aa, bár azonos, de genotípusosan, azaz genetikai összetételében eltérő. Mendel további fontos törvényei a következők: a karakterek elkülönítésének vagy szétválasztásának törvénye és a karakterek függetlenségének törvénye.
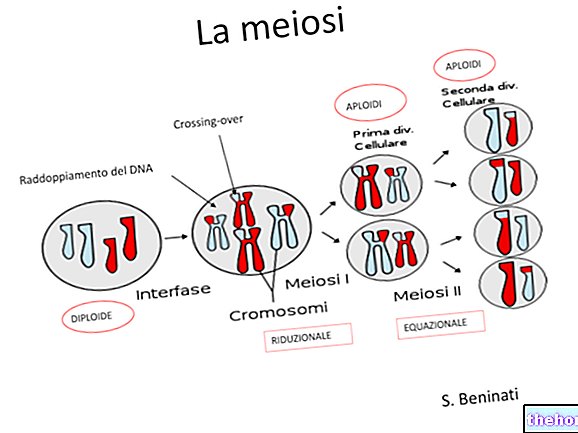
Mendel idejében a mitózis és a meiózis jelenségei még nem voltak egyértelműek, de ma már tudjuk, hogy a meiózisban az ivarsejtek minden párból csak egy kromoszómát kapnak, és hogy kizárólag megtermékenyítéssel ezek a kromoszómák véletlenszerűen visszatérnek a párzáshoz.
Ha azt gondoljuk (átmeneti egyszerűsítésképpen), hogy egy bizonyos tényező egyetlen kromoszómapáron lokalizálódik, akkor azt látjuk, hogy az eukarióta (diploid) szervezetben a faktorok párban vannak jelen, és csak az ivarsejtekben (haploid) van egyetlen ahol párban vannak jelen, lehetnek azonosak vagy különbözőek.
Ha két egyenlő tényező (akár domináns, akár recesszív, GG vagy gg) egyesült a zigótában, a kapott egyedről azt mondják, hogy homozigóta az adott karakterre, míg a heterozigóta olyan, amelyben két különböző tényező egyesült (Gg).
Az alternatív tényezőket, amelyek meghatározzák az egyén jellegét, alléloknak nevezzük, esetünkben G és g a domináns allél és a borsó színkarakterének recesszív allélje.
Egy bizonyos karakter allélje kettőnél is több lehet. Ezért beszélünk diallelikus és poliallelikus karakterekről, illetve genetikai dimorfizmusról és polimorfizmusról.
Megállapodás szerint a kísérleti kereszt generációit P, F1 és F2 szimbólumokkal jelölik, amelyek jelentése:
P = szülői generáció;
F1 = első gyermekkori generáció;
F2 = második generációs fióka.
A mendeli keresztben a sárga X zöld az összes sárgát adja; az utóbbiak közül bármelyik kettő, egymással keresztezve, minden harmadik sárgához egy -egy zöldet ad. mindig egyenlő ivarsejteket adnak, így utódaik is egyenlőek, minden heterozigóta Mivel a sárga dominál a zöld felett, a heterozigóták mind sárgák (F1).
Ha azonban kettőt keresztezünk ezekből a heterozigótákból, azt látjuk, hogy mindegyik egyenlő valószínűséggel adhat egyik vagy másik típusú ivarsejtet. A zigótákban az ivarsejtek egyesülése is azonos valószínűséggel rendelkezik (kivéve a különleges eseteket), így az F2 -ben a négy lehetséges típusú zigóta azonos valószínűséggel jön létre: GG = homozigóta, sárga; Gg = heterozigóta, sárga; gG = heterozigóta, sárga; gg = homozigóta, zöld.
A sárga és a zöld tehát 3: 1 arányban van az F2 -ben, mivel a sárga amúgy is előfordul, amíg jelen van, míg a zöld csak sárga hiányában.
A jelenség molekuláris biológiai szempontból való jobb megértéséhez elegendő feltételezni, hogy egy bizonyos alapanyagot, a zöldet nem módosítja a g allél által termelt enzim, míg a G allél olyan enzimet termel, amely a zöldet átalakítja Ha a G allél nincs jelen a gént hordozó két homológ kromoszóma egyikében sem, a borsó zöld marad.
Az a tény, hogy a sárgaborsót két különböző genetikai struktúrával, a homozigóta GG -vel és a heterozigóta Gg -vel lehet jellemezni, lehetőséget ad a fenotípus és a genotípus meghatározására.
A szervezet genetikai tulajdonságainak (amit látunk) külső megnyilvánulását, amit a környezeti hatások többé -kevésbé módosítanak, fenotípusnak nevezzük, egyedül a genetikai jellemzők halmazát, amely a fenotípusban megnyilvánulhat vagy nem, genotípusnak nevezzük.
Az F2 sárgaborsó fenotípusa azonos, de változó genotípusú. Valójában 2/3 heterozigóta (a recesszív tulajdonság hordozói) és 1/3 homozigóta.
Ehelyett például a zöldborsóban a genotípus és a fenotípus kölcsönösen változatlanok.
Amint látni fogjuk, az F1 -ben csak az egyik szülői karakter megjelenése, és az F2 -ben mindkét karakter 3: 1 arányban való megjelenése általános jellegű jelenségek, amelyek Mendel 1. és 2. törvényének tárgyát képezik. Mindez arra utal, hogy kereszteződnek az egyedek, akik egyetlen allélpár, egyetlen genetikai karakter tekintetében különböznek egymástól.
Ha bármilyen más ilyen keresztezést hajtanak végre, a mendeli minta megismétlődik; például ha a borsót ráncos maggal és sima maggal keresztezzük, amelyben a sima allél domináns, akkor LL X 11 lesz P -ben, minden LI (heterozigóta, sima) az F1 -ben, és három sima minden ráncos F2 -ben (25 % LL, 50% LI, 25% 11). De ha most keresztezzük a homozigóta párosokat, vagyis azokat a fajtákat, amelyek több karakterrel különböznek egymástól (például GGLL, sárga és sima, ggll, zöld és regosi), akkor azt látjuk, hogy az F1 -ben minden heterozigóta lesz, mindkét domináns karakterrel, fenotipizálva, de az F2 -ben ott lesz a négy lehetséges fenotípusos kombináció 9: 3: 3: 1 számarányban, amely a lehetséges lehetséges kombinációknak megfelelő 16 lehetséges genotípusból származik négyféle ivarsejt (a zigótákban párokból kettőbe szedve).
Nyilvánvaló, hogy az első generációban együtt élő két karakter egymástól függetlenül elkülöníti egymást a harmadikban. A homológ kromoszómapárok mindegyike egymástól függetlenül szegregálódik a meiózisban, és ezt állapítja meg Mendel 3. törvénye.
Lássuk most összességében Mendel három törvényének megfogalmazását:
1a: dominancia törvénye. Ha egy allélpárt adunk, ha a megfelelő homozigóták keresztezésének utódainak csak az egyik szülői karaktere van a fenotípusban, akkor ezt dominánsnak, a másikat recesszívnek nevezzük.
2a: a szegregáció törvénye. Az F1 hibridek keresztezése három dominanciát eredményez minden recesszív esetében. A fenotípus arány tehát 3: 1, míg a genotípus arány 1: 2: 1 (25% domináns homozigóta, 50% heterozigóta, 25% recesszív homozigóta).
Amikor egynél több allélpárral különbözõ egyéneket kereszteznek, mindegyik pár az elsõ és a második törvény szerint leszármazókká válik, a többitõl függetlenül.
Ezt a három törvényt, bár Mendel nem megfelelően fogalmazta meg, elismerik az eukarióta genetika alapjaként. Mint mindig a biológia nagy elveiben, e törvények általános jellege nem jelenti azt, hogy nincsenek kivételek.
Valójában annyi lehetséges kivétel létezik, hogy ma már szokás a genetikát Mendeli és Neomendeliánusokra osztani, beleértve az utóbbiakat is, minden olyan jelenséget, amely nem tartozik a mendeli törvények hatálya alá.
Míg azonban az első kivételek kétségbe vonják Mendel felfedezéseinek érvényességét, utólag be lehetett bizonyítani, hogy törvényei általános hatályúak, de a mögöttes jelenségek számos más jelenséggel kombinálódnak, amelyek ezeket módosítják.
FOLYTATÁS: Jósolja meg gyermeke vércsoportját "













.jpg)
--dimetindene.jpg)







